 Andreas R. Hassl
Andreas R. Hassl
VI.0 Die Systematik des Taxon „Zecken”
Auf dieser in den Fachbereich der Allgemeinen Zeckenkunde einführenden Internet-Seite wird ein gestraffter Überblick über die Einordnung dieser Tiergruppe in ein zoologisches System dargelegt und so soll das Wissenswerteste zur Klassifizierung und Kategorisierung dieser Tiergruppe allgemeinverständlich veranschaulicht werden. Unter dieser Thematik subsumiert sind Daten zu der Zecken Klassifikation, zu ihrer Systematik, Taxonomie und Nomenklatur, zur adspektorischen Unterscheidung der beiden Hauptgruppen und zu ihrer Artenvielfalt in Form von Listen gültiger Artnamen.
Inhaltsverzeichnis:
- VI.1Allgemeines zur Systematik
- VI.2Das System bis zur Klasse
- ❍Die Klasse Arachnida
- VI.3Das System bis zur Ordnung
- ❍Die Ordnung Acari
- VI.4Das Taxon Ixodida
- ❍Die Familie Nuttalliellidae
- ❍Die Familie Deinocrotonidae
- VI.5Die Unterschiede zwischen Schild- und Lederzecken
- ❍Die Familie Argasidae
- ❍Eine Liste der validen Artnamen aller Lederzecken
- ❍Die Familie Ixodidae
- ❍Eine Liste der validen Artnamen aller Schildzecken
VI.1 Allgemeines zur Systematik
Eine „Zecke” ist ein Begriff, eine semantische Einheit, in dem viele, sich unterscheidende Tiere mit verschiedenem Aussehen und physiologischen Zuständen enthalten sind: Kleine und große Tiere, angesogene und hungrige, Männchen, Weibchen und Jungtiere, Tiere mit Augen und solche ohne, prächtig gefärbte, bunte und einfarbig schwarzbraune Tiere, also offensichtlich verschiedenartige „Zecken”. In der Zeckenkunde werden dafür dann jeweils eigene Fachbegriffe benutzt, also zB für eine bestimmte, heimische, den Menschen als Blutspender liebende, augenlose, schwarzbraune Zecke der Name Ixodes ricinus, der dem ⇒ biologischen Artbegriff entspricht oder aber die in Ostösterreich gebräuchlichere Bezeichnung „Gemeiner Holzbock”. Die Festlegung des Inhalts eines Begriffs und damit die schiere Behauptung der Existenz eines Tieres folgen historisch geprägten Regeln. Diese Regeln zumindest in den Grundzügen zu verstehen und anwenden zu können ist eine unbedingte Voraussetzung für eine lohnende Beschäftigung mit biologischen und parasitologischen Themen.
Die Benennung (Namensgebung) und Identifizierung (Begriffsbildung) von belebten und unbelebten Dingen ist ein dem Menschen immanenter geistiger Prozess, weil unser Denken auf dem Manipulieren hierarchisch angeordneter Begriffe beruht. Im abendländischen Kulturkreis wird in diesem Zusammenhang häufig das Bild von Adam im Paradies bemüht, dessen vorzügliche Aufgabe die Benennung der Mitgeschöpfe als Fundament seiner Herrschaft über sie war (1. Moses 1,28: „Gott sprach zu ihnen: . . . und herrscht über die Fische des Meeres, über die Vögel des Himmels und über alle Tiere, die sich auf dem Land regen.”). Einteilungen in Klassen und Hierarchiebildung ist daher in der Biologie so alt wie diese selbst. Die naturwissenschaftliche Beschäftigung damit wird Systematik genannt.
Die klassische Systematik beschäftigt sich daher mit der Einteilung (= Taxonomie), Benennung (= Nomenklatur, eine Kunstfertigkeit) und Identifizierung (= Diagnostik) der Lebewesen. Die Identifizierung eines Parasiten ist daher ein Teilgebiet der Systematik. Eine treffsichere Diagnostik ist ein integraler Bestandteil der heilenden Medizin sein. Eine "moderne" Systematik umfasst zudem auch noch die Rekonstruktion der Stammesgeschichte der Organismen (Phylogenie) und die Erforschung der Prozesse, die zu der gegenwärtigen Vielfalt von Lebewesen führten (Evolutionsbiologie).
Die vermutlich erste überlieferte, abendländische Systematik stammt von Aristoteles (384-322 vChr), der die ihm bekannten Lebewesen in einer Stufenleiter (Scala Naturae) nach dem Grad ihrer „Perfektion” anordnete. Einige seiner Bezeichnungen sind bis heute in Verwendung (zB Coleoptera, Diptera) und die Spukgestalt der „primitiven” Form eines rezent existierenden Lebewesens geistert heute noch durch das Denken mancher Biologen.
Carl von Linné verwendete in seinem grundlegenden Werk Systema Naturae [1758] eine binäre Nomenklatur zur Benennung von Tierarten. Der menschlichen Ordnung des römisch-rechtlichen Rechtssystems mit den Begriffen Gattungs- und Speziessachen wurde eine göttliche Ordnung mit „belebten” Genera und Spezies gegenübergestellt. Das Grundkonzept ist eine typologische Definition der Art, das heißt die Merkmalsfülle eines Organismus wird auf einige wenige Schlüsselmerkmale reduziert und ein Typus (= ein Schöpfungsakt) abstrahiert. Linnés Einteilung und hierarchische Ordnung der Organismenfülle in drei Reiche und folgend in Klassen, Ordnungen, Gattungen und Arten sind heute noch in Verwendung, weil dieses System eine schnelle und akkurate Erkennung eines Taxons ermöglicht.
Das Konzept einer ⇒ biologischen Art nach Ernst Walter Mayr (1904-2005) beruht auf einer objektivierbaren Feststellung: Eine Art ist eine Gruppe natürlicher Populationen, die sich untereinander kreuzen können und von anderen Gruppen reproduktiv isoliert sind. Dieses bestechende Artkonzept lässt jedoch das Phänomen der hierarchisch geordneten Begriffswelt des Menschen außen vor und ist nur an sexuell sich vermehrenden, gegenwärtig existierenden Lebewesen anwendbar.
Mit der breiten Akzeptanz der Evolutionstheorie werden diese teilweise künstlichen Systeme in ein „natürliches” System umgebaut. Letzteres soll die Abstammungsverhältnisse, die Phylogenie, besser widerspiegeln. Dabei spielte zunächst die Homologisierung von Organen eine große Rolle, später dann Ähnlichkeiten von Proteinen. Heute werden Hinweise auf den Verwandtschaftsgrad aus ⇒ morphologischen, anatomischen, biochemischen, physiologischen, zytologischen, ethologischen und zunehmend genetischen Merkmalen abgeleitet.
Die konsequent phylogenetische Systematik nach Willi Hennig (1913-1976) ist zwar wissenschaftlich besser fundiert als ihre Vorläufer und Konkurrenten, aber sehr unübersichtlich und meist auch äußerst unpraktisch. In ihr werden die Taxa (= systematische Einheiten) nur von den Arten gebildet, die eine geschlossene Abstammungsgemeinschaft, ein ⇒ Monophylum, bilden. Die kleinste, real existierende Einheit ist das ⇒ Taxon Art, was in der Konsequenz zur völlig unüberschaubaren Aufsplittung der Organismenfülle in winzige Einheiten mit jeweils einem eigenen Namen führt. Im phylogenetischen Konzept werden auf höheren Ebenen nur jene Taxa zusammengefasst, die durch Synapomorphien charakterisiert sind, und von Taxa mit ⇒ Autapomorphien unterschieden werden können. Eine Autapomorphie ist eine evolutionäre Neuheit eines Taxons, eine Synapomorphie stellt ein Merkmal dar, welches ausschließlich den direkt aus dem Stammtaxon entstandenen Arten gemein ist. Problematisch ist und bleibt die Unterscheidung zwischen Aut- und Synapomorphien, die im Zweifelsfall dann wiederum - wie im Linnéschen System - nach subjektiven Kriterien getroffen wird. So wird daraus allerdings wiederum ein typologisches Konzept mit klassischen hierarchischen Benennungen, nur diesmal ohne Gott dafür zu bemühen.
VI.2 Das System bis zur Klasse
Nachfolgend eine textliche Darstellung jenes Stranges im System der Tiere, der zu den Spinnentieren führt, inklusive einer Auflistung der jeweiligen Schwestertaxa im nächsthöheren Taxon. Obgleich es sich um das derzeit gültige, konsequent phylogenetische System nach Burda et al. [2008] handelt, sollten die bald 250-jährigen Rangbezeichnungen der Taxa und damit die zwangsläufig subjektive Gewichtung von Merkmalen keinesfalls unbeachtet bleiben:
Domäne Eukaryoten
Reich Vielzellige Tiere (Metazoa)
Taxon Gewebetiere (Eumetazoa)
Taxon Zweiseitentiere (Bilateria)
Stammgruppe Urmünder (Protostomia)
Überstamm Lophotrochozoen (Lophotrochozoa)
Überstamm Häutungstiere (Ecdysozoa)
Taxon Scalidophora
Taxon Nematoida
Taxon Panarthropoda
Stamm Bärtierchen (Tardigrada)
Stamm Stummelfüßer (Onychophora)
Stamm Gliederfüßer (⇒ Arthropoda)
Unterstamm Pancrustacea inkl. Krebstiere (Crustacea) und Sechsfüßer (Hexapoda)
Unterstamm Tausendfüßer (Myriapoda)
Unterstamm Kieferklauenträger (Chelicerata)
Klasse Asselspinnen (Pycnogonida)
Klasse Spinnentiere (Arachnida) (inkl. Skorpione)
Klasse Pfeilschwanzkrebse (Merostomata)
Die Definitionen der höheren Taxa umfassen jeweils immer auch die Eigenschaften und Merkmale, die letzten Endes der Ordnung Ixodida eigen sind. Die folgende Übersichtsdarstellung folgt in veränderter Form der Ansicht Kaestners [1993].
- Eukaryoten sind alle Lebewesen, deren Zellen einen Zellkern beinhalten. Eukaryotische Zellen sind mittels Zellorganellen strukturiert, die verschiedene Aufgaben innerhalb der Zelle erfüllen.
- Metazoa oder genauer Eumetazoa sind eukaryotische, heterotrophe, zellwandlose Lebewesen (= Tiere), deren zumindest zu Epithelien differenzierte Zellen meist sehr enge Aufgabenbereiche erfüllen.
- Zweiseitentiere oder Bilateria sind Eumetazoen, deren Körper durch eine Mediosagittalebene in zwei spiegelbildlich gleiche Hälften zerlegt werden kann. Sie besitzen zudem eine Bauch- (⇒ Ventral-) und eine Rückenseite (⇒ Dorsalseite), aber auch ein Vorder- und ein Hinterende. Der verdauende Hohlraum mündet in einen After und es sind weitere Körperhöhlen vorhanden.
- Urmünder oder Protostomia sind Bilateria mit einem vollständigen Verdauungstrakt, bei denen in der Embryonalentwicklung der Mund aus dem schlitzförmigen Urmund hervorgeht, während der ⇒ Anus sekundär die Körperoberfläche durchbricht oder aus dem hinteren Tei- undes entsteht. Das Zentralnervensystem im Rumpf bildet bei den Protostomiern ventral starke Längsstränge aus.
- Häutungstiere oder Ecdysozoa sind Protostomier, die vermittels der Sequenzierungsdaten der rRNS von den Lophotrochozoen und den Plattwurmartigen abgetrennt wurden. Das gemeinsame ⇒ morphologische Merkmal der Häutungstiere ist eine dreilagige, aus organischem Material aufgebaute Cuticula; ein weiteres Merkmal ist die Häutung, die durch Hormone ausgelöst wird.
- Panarthropoda sind Ecdysozoa mit den gemeinsamen Merkmalen Besitz von Beinen mit einem krallenbesetzten Fuß, Besitz eines Bauchmarks und eine ⇒ Segmentierung des Körpers.
- Gliederfüßer oder Arthropoda sind Panarthropoda, deren Körper mit einer Chitinkutikula überzogen ist und deren Segmente zumindest im vorderen Drittel des Körpers paarige Extremitäten tragen. Diese bestehen aus mit einem Gelenk miteinander verbundenen, starren Chitinröhren. Wenigstens eines dieser Gliedmaßenpaare ist zu einem Fühler oder einem Mundwerkzeug differenziert.
- Kieferklauenträger oder Chelicerata sind Arthropoda, die sich durch den Besitz einer spezifisch umgewandelten Extremität auf dem ersten Kopfsegment auszeichnen. Diese Mundwerkzeuge heißen Chelizeren und sind die ersten Extremitäten des Körpers. Antennen, die ersten Extremitäten der Pancrustacea und der Myriapoda fehlen.
- Spinnentiere oder Arachnida sind luftatmende Chelicerata, deren Leib in zwei Abschnitte gegliedert ist: Der aus dem Akron und sechs ⇒ Segmenten gebildete Vorderleib (= Prosoma, im Falle der Zecken Capitulum genannt) und der Hinterleib (= Opisthosoma). Der Hinterleib trägt keine voll ausgebildeten, sondern höchstens rudimentäre Extremitäten.
Die Klasse Arachnida (Spinnentiere)
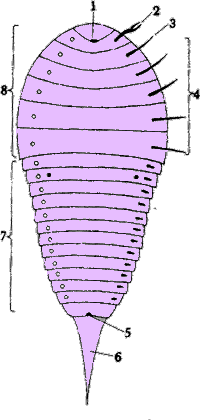
Abb. VI.1: Schema eines Spinnentiers. cit. Grzimek [1980]. Abbildungslegende: 1: Mund; 2: Chelizere; 3: Pedipalpus; 4: Laufbeine; 5: After; 6: Schwanzstachel; 7: Hinterkörper; 8: Vorderkörper.
Aufgrund molekularbiologischer Daten werden heute die Gliederfüßer (⇒ Arthropoda) mit ihrer Teilmenge der Arachnida mit den Cycloneuraliern, das sind die Nematoda (Fadenwürmer), Nematomorpha (Saitenwürmer), Loricifera (Korsetttierchen), Priapulida (Priapswürmer) und die Kinorhyncha (Hakenrüssler), zu den Ecdysozoa (Häutungstiere) zusammengefasst. Gemeinsamkeiten, dh synapomorphe Merkmale, der Cycloneuralia mit den Arthropoda sind:
- Die dreischichtige Alpha-Chitin-Cuticula, die durch regelmäßiges Häuten gewechselt werden muss, um dabei das Körpervolumen zu vergrößern.
- Die Häutungen werden durch ein Steroidhormon (Ecdyson) induziert.
- Das Abhandenkommen von äußeren lokomotorischen Zilien.
- Das Vorhandensein eines Mixocoel und das damit in Verbindung stehende, offene Blutgefäßsystem.
- Und, als sehr umstrittene Gemeinsamkeit, die Befähigung, eine Infektion mit Bakterien der Gattung Wolbachia erleiden zu können.
Historisch bedeutsam, auf Cuvier [1795] zurückgehend ist die Ansicht, dass die Ringelwürmer (Annelida) und die Gliederfüßer Schwestergruppen sind. Die wichtigsten Synapomorphien der Arthropoden und der Anneliden sind danach:
- Ein Strickleiternervensystem mit einem in Bewegungsrichtung vorne gelegenen Zerebralganglion und einem Paar Ganglien mit ventraler Kommissur in jedem Segment.
- Die Zusammensetzung des Körpers aus einem Vorderteil (Acron), einem Hinterteil (Pygidium) und einer nur theoretisch bestimmbaren Anzahl ursprünglich gleichartig gebauter (= homonomer) Abschnitte (= Segmente).
- Die ursprünglich identische Anatomie dieser Segmente, die alle ein Paar Coelomsäcke, ein Paar Metanephridien, ein Paar Ganglien und ein Paar Extremitäten besaßen.
- Eine Kutikula mit Borstenhaaren (Setae) aus Chitin.
Die Arachnida Lamarck, 1801 oder Spinnentiere (Arachnida; von altgriechisch αραχνη aráchne = Spinne) sind eine Klasse der Gliederfüßer (Arthropoda). Gemeinsam mit den Pfeilschwanzkrebsen bilden sie den Unterstamm der Kieferklauenträger (Chelicerata). Ihr Körper war ursprünglich in zwei Teile gegliedert, in das Prosoma (Vorderleib) und das Opisthosoma (Hinterleib). Im Falle der Zecken un der anderen Milben sind die beiden Körperteile miteinander verschmolzen und adspektorisch nicht differenzierbar. Ein auffälliges Merkmal sind ihre vier Paar Laufbeine der Adulttiere. Dieses Merkmal und ihr zweigeteilter Körper unterscheidet sie von anderen Ordnungen innerhalb der Klasse, insbesondere von den Insekten. Allerdings haben die Larven vieler Milbenarten ebenso wie die Insekten nur sechs Beine. Andere Extremitäten sind im Laufe der Evolution zu Werkzeugen wie Giftklauen, Scheren, Tastern oder Mundwerkzeugen umgebildet worden. Die meisten Spinnentiere leben räuberisch, die Zecken sind ⇒ obligatorisch blutsaugend. Wie alle Gliederfüßer besitzen Spinnentiere ein Strickleiternervensystem. Falls Augen vorhanden sind, sind es mehrere Punktaugen. Einige Arten haben sehr leistungsstarke Augen entwickelt, dies trifft allerdings vermutlich nicht auf die Tiere in der Gruppe Zecken zu.
VI.3 Das System bis zur Ordnung
In die Klasse der Spinnentiere reihen sich derzeit folgende Ordnungen:
REM: Das Kreuz kennzeichnet eine ausgestorbene Ordnung, die Buchstaben am Ende jeder Kurzbeschreibung beziehen sich auf die Abbildung 2 unterhalb der Liste.
- Amblypygi Thorell, 1883, dt: Geißelspinnen: Nicht zu verwechseln mit den Geißelskorpionen. Weltweit sind etwas mehr als 200 Arten dieser Tiere bekannt, in 5 Familien und 17 Gattungen. Diese Tiere erreichen eine Körpergröße von 10 bis 45 mm. Fast alle Geißelspinnen leben in den Tropen und Subtropen, die meisten in Regenwäldern. Im Mittelmeerraum kommen zwei Arten vor, eine davon auf zwei griechischen Inseln. Die Palpen sind zu Fangapparate umgestaltet. Die Beine des ersten Laufbeinpaares sind sehr lang und dienen als Tastorgane; sie haben eine höhere Anzahl ursprünglicher Beinglieder behalten als die anderen Beine. Als effektive Laufbeine dienen nur die folgenden drei Paare. Wehr- oder Giftdrüsen fehlen den Geißelspinnen. (E)
- Araneae Clerck, 1757, dt: Webspinnen: Über 46 000 Arten in 114 Familien. Allgemein bekannte Tiere, die durch den Besitz von Spinndrüsen gekennzeichnet sind. Webspinnen leben zumeist räuberisch und ernähren sich von Gliedertieren, insbesondere von Insekten. Fast alle Webspinnen sind Landtiere. Sie sind kosmopolitisch verbreitet, doch findet man in den heißen Zonen eine höhere Artenzahl und größere Arten. (A)
- Haptopoda✟
- Opiliones Sundevall, 1833, dt: Weberknechte, Schneider, Kanker: Tiere mit einer Körperlänge von 2 bis 22 Millimeter und einem einteiligen, meist gedrungenem Körper und langen, dünnen, beweglichen Beinen. Das Unterscheidungsmerkmal zu den Webspinnen ist die Verwachsung des Vorder- und des Hinterkörpers; der Hinterkörper ist aber stets gegliedert. Weberknechte verfügen weder über Spinndrüsen noch über Giftdrüsen, aber über Stinkdrüsen. Die Männchen haben einen Penis. Sie ernähren sich hauptsächlich von kleinen Gliederfüßern und von toten Insekten. Bevorzugte ⇒ Habitate sind Laubwälder, Gärten, Wiesen, Hecken und naturnahe Parks. Etwa 6 500 Arten sind in mehr als neun Familien gegliedert und die Ordnung ist weltweit verbreitet. (G)
- Palpigradi Thorell, 1888, dt: Palpenläufer: Die Tiere sehen den Geißelskorpionen änlich. Die Palpenläufer sind vollkommen farblos und ihnen fehlen Augen und Fächerlungen. Möglicherweise gibt es in dieser Gruppe relativ häufig ⇒ parthenogenetische Formen. Weltweit sind knapp 80 Arten bekannt, die Körperlängen von nur zwei bis drei Millimetern erreichen. Aus Europa sind 26 Arten bekannt. (H)
- Phalangiotarbida✟
- Pseudoscorpiones de Geer, 1778, dt: Pseudo- , Bücher- oder Afterskorpione: Diese Tiere sehen den Skorpionen ähnlich, allerdings mit Körperlängen von nur zwei bis sieben Millimetern. Ihnen fehlt aber der Stachel der Skorpione, sie besitzen ein abgerundetes Hinterende. Pseudoskorpione finden sich vor allem am Boden in der Laubauflage sowie in Moospolstern und Pilzmatten, aber auch häufig unter loser Baumrinde. Sie kommen in beinahe allen Landlebensräumen vor, inklusive der Meeresküste sowie den menschlichen Behausungen. Einige Spezies lassen sich von Fliegen, Käfern oder anderen Insekten transportieren (Phoresie; Transportparasitismus). Sie sind räuberisch und leben von anderen, kleineren Gliederfüßern wie Springschwänzen. Weltweit sind etwa 3 000 Arten bekannt und ungefähr 100 Arten werden in ⇒ Zentraleuropa regelmäßig gefunden. (C)
- Ricinulei Thorell, 1876, dt: Kapuzenspinnen. Weltweit existieren etwa 40 Arten, die die tropischen Wälder Afrikas und Südamerikas bewohnen. Sie sehen zwar Webspinnen ähnlich, man erkennt sie aber relativ leicht an dem langen zweiten Laufbeinpaar, das zu einem Tastorgan umgebildet wurde und an den vier, einziehbaren Segmenten des Hinterkörpers. (J)
- Schizomida Petrunkevitch, 1945, dt: Zwerggeißelskorpione. Ca. 230 Arten weltweit in zwei rezent existierenden Familien. Die Körperlänge beträgt maximal 18 mm. Sie leben vor allem im Boden und in Höhlen. Die Vorderkörpersegmente besitzen jeweils eigene Rückenplatten. Den Tieren fehlen die Augen. Die Palpen sind schlank und ohne Schere, das Flagellum kurz und beim Männchen knopfartig verdickt. Die Zwerggeißelskorpione sind in allen tropischen Regionen der Erde heimisch, es gibt auch eine Art, die in ⇒ Zentraleuropa als ⇒ Neozoon auftritt.
- Scorpiones C.L. Koch, 1837, dt: Skorpione: Allgemein bekannte und mit einer magischen Aura umgebene Tiere von 9 mm bis 21 cm Körperlänge. Sie zeichnen sich durch bewegliche Finger an den Palpen, die zu Scheren umgebildet wurden, und einem Giftstachel am hinteren Ende aus. Sie leben vorwiegend auf und in sandigen oder steinigen Böden der tropischen und subtropischen Wüsten und Halbwüsten. Ca. 1400 Arten in 13 Familien. (D)
- Solifugae Sundevall, 1833, dt: Walzenspinnen: Mehr als 1 000 bekannte Arten in 12 Familien und 153 Gattungen. Die Tiere messen vom Kopf bis zum Hinterleib bis zu 70 mm. Charakteristisch ist der deutlich gegliederte, langgestreckte, meist in seiner ganzen Breite dem gegliederten Vorderleib angewachsene Hinterleib aus elf Segmenten. Weitere Kennzeichen sind die sehr großen, am Grund blasig aufgetriebenen und scherenförmigen Chelizeren und die senkrecht stehenden Palpen. Die Walzenspinnen leben in den Trockengebieten, vor allem in Wüsten und Steppen aller Kontinente außer der Antarktis und Australiens. Einige Arten sind im Mittelmeerraum zu finden. Die meisten Arten sind nacht- oder dämmerungsaktiv. Walzenspinnen sind räuberisch, sie fressen Insekten, Webspinnen, Skorpione, anderen Walzenspinnen und sogar Reptilien. (F)
- Thelyphonida O. P.Cambridge, 1872, dt: Geißelskorpione: Das hintere Körperende ist spitz ausgezogen und formt eine Geißel. Die Körperlänge ohne Geißel liegt zwischen 25 und 85 mm. Das erste Laufbeinpaar ist zu einem langen Tastorganen mit einer vermehrten Anzahl an Beingliedern umgestaltet. Die Palpen sind mit kleinen Scheren ausgestattet oder scherenlos. Die Tiere sind räuberisch, sie fressen Insekten, Skorpione, Tausendfüssler, Isopoden und Schnecken. Mehr als 180 Arten in 4 Familien. Sie leben in den R- l;ldern der Tropen und Subtropen Asiens und der beiden Amerikas. Aus Afrika ist nur eine Art bekannt. (I)
- Trigonotarbida✟
- Acari oder Acarina Leach, 1817, dt: Milben: Diese Ordnung wird im nächsten Abschnitt genauer dargelegt. (B)

Abb. VI.2: Die Abbildung zeigt das typische Aussehen von einigen Vertretern der Ordnung.
A: Araneae dt: Spinnen; B: Acari dt: Milben; C: Pseudoscorpiones dt: Pseudoskorpione; D: Scorpiones dt: Skorpione; E: Amblypygi dt: Geißelspinnen; F: Solifugae dt: Walzenspinnen; G: Opiliones dt: Weberknechte; H: Palpigradi dt: Palpenläufer; I: Uropygi dt: Geißelskorpione; J: Ricinulei dt: Kapuzenspinnen.
Die Ordnung Acari Leach, 1817, dt: Milben
Die Milben sind mit mehr als 50 000 beschriebene Arten und wahrscheinlich mehr als einer Million Arten in 546 Familien die größte Untergruppe der Arachnida. Dieses Taxon wird häufig als Unterklasse bezeichnet. Es ist vermutlich monophyletisch vor 410 Millionen Jahren entstanden. cit. Mans et al. [2016]. Die Milben zeichnen sich durch eine geringe Größe - 0,08 bis 1 Millimeter, die größten Milben sind Zecken mit bis zu 20 mm - aus, durch einen ungeteilten Körper, und oft durch eine starke Reduktion verschiedener Körperteile. Die Mundwerkzeuge sind umgewandelte Beine, sie sind mit ihren Basen stets zu einem ⇒ Capitulum verschmolzen. Die ⇒ Genitalöffnung auf der Bauchseite ist nach vorne verschoben und liegt zwischen den Beinen. Das Erscheinungsbild der einzelnen Arten ist sehr unterschiedlich. Für einen genauen Vergleich des Aufbaus von Milben mit dem der anderen Arachnida vergleiche Kaestner [1993].
Milben leben praktisch überall auf der Erde, auf dem Land wie im Wasser, selbst in kalten Wüsten wie in der Antarktis gibt es Milben. Die nächsten Verwandten unter den Arachniden scheinen die Weberknechte und die Kapuzenspinnen zu sein, mit denen die Milben eine Anzahl von Merkmalen gemein haben. Die Phylogenie sowohl innerhalb der Gruppe als auch die der Klasse ist allerdings nicht gut untersucht. Fest zu stehen scheint aber, dass sich die Acari sehr früh in zwei Entwicklungslinien aufgeteilt haben, die sich heute im Range von Ordnungen präsentieren, wenn die Milben als Unterklasse gesehen werden:
- Die ursprünglich nicht parasitischen Anactinotrichida oder Parasitiformes, inklusive der manchmal als dritte Gruppe betrachteten Opilioacariformes, und
- die Actinotrichida (oder Acariformes).
Die Unterschiede zwischen diesen beiden Gruppen können im Falle von Interesse dem basalen Lehrbuch jedes Zoologie-Studiums Kaestner [1993] entnommen werden. Die Namen beziehen sich auf die chemische Zusammensetzung der Körperborsten: Denen der Anactinotrichida fehlt das doppelbrechende Actinopilin. Die Anactinotrichida teilt man in vier Gruppen, die Actinotrichida in zwei. Diese haben meist den Rang von Unterordnungen oder wiederum von Ordnungen.
- 01. Anactinotrichida
- 01.01. Opilioacarida With, 1902. Purpurrote, violette oder grünlichblaue, als ursprünglich angesehene, große Milben. Verbreitet in den wärmeren Gegenden der Welt, auch in Europa, aber selten. Mit einem ovalen bis länglichen, relativ weichen Körper. Zehn Gattungen in einer Familie.
- 01.02. Holothyrida Thon, 1905. 2 bis 7 mm große Milben mit einem ovalen, hellroten bis dunkelbraunen, stark sklerotisierten Körper. Die Tiere leben meist in der Humusschicht tropischer Wälder. Sie ernähren sich von der Hämolymphe toter Arthropoden. 25 beschriebene Arten in 10 Gattungen. Es ist die Schwestergruppe der Ixodida, da sie mit diesen gemeinsam das Merkmal des Haller'schen Organs besitzen.
- 01.03. Mesostigmata Canestrini, 1891 = Gamasida, dt: Raubmilben. 0,2 bis 2 mm große, stark bepanzerte braune oder rötliche bis braune Milben, meist frei-lebend und räuberisch. Einige Arten parasitieren auf anderen Arthropoden. Sie können an dem einzelnen Paar an Tracheenöffnungen erkannt werden, die ⇒ lateral am Körper liegen. Die Raubmilben sind eine sehr große Gruppe: 100 Families, 900 Gattungen und mehr als 11 500 Arten.
- 01.04. Ixodida Leach, 1815 = Metastigmata, dt: Zecken. Die Zecken unterscheiden sich von den anderen Gruppen durch zwei ⇒ autapomorphe Merkmale, eines Luftfeuchte absorbierenden Azinarsystems (cit. Mans et al. [2016]), und das Gené'sche Organ, und auch durch ihre Körpergröße als Adulte. Sie besitzen ebenso wie die Holothyrida ein Haller'sches Organ. Es wird angenommen, dass die ersten Ixodida Gewebsflüssigkeits-Sauger an Wunden von Wirbeltieren waren.
- 02. Actinotrichida
- 02.01. Trombidiformes Reuter, 1909. Die Trombidiformes sind eine große, sehr heterogene Milbenordnung mit etwa 22 000 Arten in 125 Familien. Die Gruppe weist nur wenige Synapomorphien auf. Viele Arten haben eine auffallende Körperfärbung, frei-lebende Arten sind oft rot. Der Körper ist meist nur schwach sklerotisiert. Die Milben leben meist fungivor, allerdings beherbergt dieses ⇒ Taxon auch medizinisch wichtige Milben (zB die Demodikose-Erreger) und Landwirtschaftsschädlinge.
- 0202. Sarcoptiformes Reuter, 1909. Milben mit einer Körperlänge von 0,15 mm bis 5 mm. Sarcoptiformes besitzen weder ursprüngliche Stigmenöffnungen noch Peritremen; sekundäre Atmungsöffnungen sind manchmal vorhanden. Das ⇒ Gnathosoma ist frei liegend oder in eine schachtelartige Struktur, das Camerostoma, zurückgezogen. Die Basen der Chelizeren sind nie von einem sklerotisierten Ring bedeckt, die Chelizeren selbst sind zweigliedrig. Die Palpen haben ein bis fünf freie Segmente. Die intercoxale Region ist ohne sternale oder genitale Schildelemente. Leben von organischem Material (saprovor oder parasitisch) und sind deshalb oft mit größeren Tieren assoziiert. Ca. 15 000 beschriebene Arten in etwa 230 Familien. Dieses Taxon beinhaltet derzeit zwei Untergruppen:

Abb. VI.3: Die Obstbaumspinnmilbe Panonychus ulmi, ein Vertreter der Trombidiformidae.

Abb. VI.4: Die Abbildung zeigt zwei Vertreter der Oribatida: links Damaeus onustus Koch, 1841, rechts Euzetes globulus Nicolet, 1855.
- 02.02.1. Astigmata Canestrini, 1891. Circa 80 Familien mit 1 000 Genera (sic!) und 3 400 Arten. Viele parasitische Arten wie zB die Federmilben, der Krätze- und die Räudeerreger.
- 02.02.2. Oribatida Dugés, 1833, dt: Horn- oder Moosmilben. Die Hornmilben sind eine Ordnung der Milben, die überwiegend im Humus und Boden, aber auch in feuchtem Moos und selten im Süßwasser leben. Fossilien belegen, dass diese Tiergruppe erstmals vor etwa 380 Mill Jahren im Devon auftauchte. Dieses ⇒ Taxon ist äußerst arten- und formenreich. Heute sind weltweit etwa 10 000 Hornmilben-Arten bekannt, von denen ca 60 Familien mit 400 Arten in ⇒ Zentraleuropa vorkommen. Die meisten Arten sind stark gepanzert, braun bis schwarz gefärbt, und haben eine kugelige Form und eine Körperlänge von 0,2 bis 1,4 mm. Hornmilben fressen Pflanzenteile wie zum Beispiel Laubblätter, seltener leben sie räuberisch oder als Aasfresser, nie jedoch parasitisch. Als Bodenbildner ist ihre Anzahl im Boden bedeutend, ihre Individuendichte kann auf einem Quadratmeter Waldboden bis zu 50 000 Tiere betragen, was einem Lebendgewicht von bis zu 20 kg je Hektar entspricht. Einige Hornmilben sind Zwischenwirte von Bandwürmern, die an Haus- oder Wildtieren parasitieren und von denen einige wenige in seltenen Fällen auch den Menschen befallen können (Moniezia expansa Rudolphi, 1810, Bertiella studeri Blanchard, 1891, Bertiella mucronata Meyner, 1895).
VI.4 Das Taxon Ixodida Leach, 1815, Zecken, ticks, tique, garrapatas
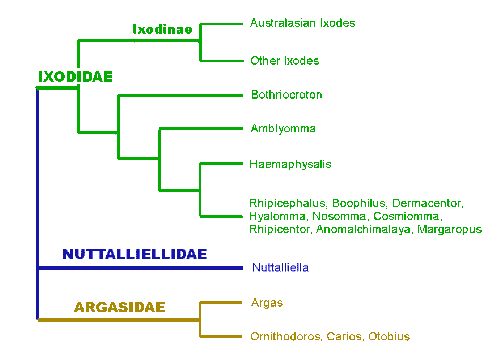
Abb. VI.5: Ein einfacher phylogenetischer Baum der Zeckengattungen. cit. Barker & Murrell [2004]
Die Zecken mit ihren derzeit 954 validen Arten in 21 Gattungen ist unter den Milben sicherlich die am besten bekundete und erforschte Gruppe, und auch diejenige, die schon am längsten bekannt ist. Zudem lässt sich diese Gruppe durch den Besitz von wahrscheinlich sogar zwei ⇒ autapomorphen Merkmalen, das Luftfeuchte absorbierende Azinarsystems und das Gené'sche Organ, sehr gut von den anderen Milben abtrennen. Trotzdem ist die Stellung dieser Gruppe im System immer noch in Diskussion, weshalb die Gruppe hier in vorsichtiger Weise als ⇒ Taxon bezeichnet wird. Gegenwärtig werden die Zecken als eigene Ordnung aufgefasst, die in die Überordnung Parasitiformes eingereiht wird.
Die Zeckenfauna der Welt besteht eben aus derzeit 954 Arten in vier Familien, den Ixodidae dt: Schildzecken, den Argasidae dt: Lederzecken, den Nuttalliellidae und den ausgestorbenen Deinocrotonidae✟. Die letzten beiden Familien tragen keine deutschen Namen. Es besteht eine Mindermeinung, dass die Nuttallielliden zwischen den Holothyrida und den Mesostigmata stehen, was bedeutet, dass sie die Fähigkeit zum Blutsaugen unabhängig von den Ixodiden erworben haben. cit. Mans et al. [2016]. Die Zecken sind derzeit auf 21 Gattungen aufgeteilt, die Anzahl der Gattungen hängt von der taxonomischen Ansicht des Bearbeiters ab. Rechter Hand sieht man einen als "working hypothesis" bezeichneten, phylogenetischen Baum der Zeckengattungen, der allerdings stark vereinfachend und der sicher nicht mehr den aktuellen Wissenstand repräsentierend ist. cit. Barker & Murrell [2004].
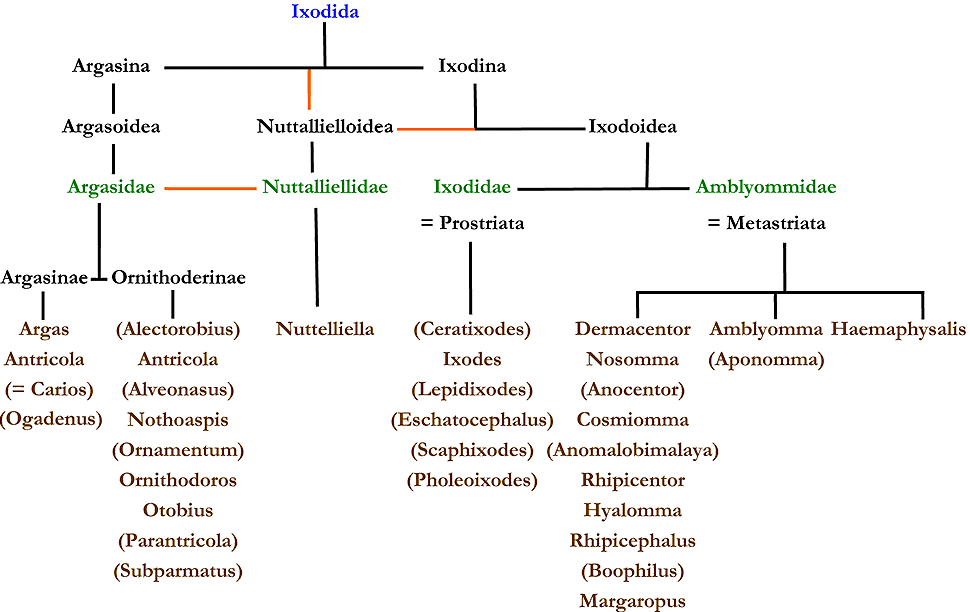
Abb. VI.6: Die systematische Position von Zecken-Taxa. cit. Camicas & Morel [1977].
Linker Hand sieht man einen deutlich mehr differenzierenden Baum der systematischen Position von den meisten 1977 anerkannten Zeckentaxa. In modifizierter Form cit. Camicas & Morel [1977]. Einige Gattungen sind nach jüngeren Meinungen inzwischen obsolet oder zu Subgenera geworden, andere, insbesondere fossile, dazu gekommen. Diese sind hier nicht aufgeführt. Die Ordnung ist in blauer Schrift dargestellt, die Familien in grüner, die Gattungen in brauner. Obsolete Gattungen in Klammern, Haupt- und meist auch Sammelgattung (für die obsoleten Taxa) in fetter Schrift. Orange Balken bedeuten eine bis heute ungeklärte systematische Position.
Die Familie Nuttalliellidae Schulze, 1935

Abb. VI.7: Habitus von Nuttalliella namaqua.
© verändert: //en.wikipedia.org/wiki/File:Nuttalliella_namaqua.png.
Die Familie Nuttalliellidae ist monotypisch: Die einzige bisher bekannt gewordene Art Nuttalliella namaqua Bedford, 1931 lebt in Südafrika, in der Limpopo- und der Northern Cape Provinz, in Südwestafrika, Namibia, und in Tansania. Die Weibchen wurden in den 20-er Jahren des 20. Jahrhunderts in den semiariden Gebieten des Namaqualandes und der Kapprovinz auf nicht beschriebenen Wirten gefunden, in Tanzania auf einem kleinen Fleischfresser, einem Nagetier und im Nest der Maidschwalbe, Cecropis abyssinica (Guérin-Méneville, 1843). Derzeit gilt diese Art allerdings als Generalist, der als Nymphe und Weibchen bevorzugt an Reptilien parasitiert, insbesondere an Skinken, Geckos und Gürtelechsen. cit. Mans et al. [2014]. Die Larven saugen hauptsächlich Blut an mäuseartigen Nagetieren. Die meisten Larven wurden an verschiedenen Arten von Mäusen (Rodentia: Muridae) gefunden, einige wenige Larven wurden von Felsenratten (Rodentia: Petromuridae), Hundeartigen (Carnivora: Canidae), Hasen (Lagomorpha: Leporidae) und Elefantenspitzmäusen (Macroscelidea: Macroscelididae) abgesammelt. cit. Apanaskevich [2021]. Besonders erwähnt werden die Felsenratten-Arten Aethomys namaquensis (A. Smith, 1834), Aethomys chrysophilus (de Winton, 1897) und die Stachelmaus Acomys spinosissimus Peters, 1852. Es könnte sich also um eine ⇒ telotropische Zecke handeln oder aber um eine Art, deren Generalistentum durch zahllose Wirtswechsel im Laufe von 300 Mill Jahren erzwungenen worden ist. Auch die Männchen und die Subadulten sind inzwischen entdeckt worden und Individuen dieser Art im Labor aufgezogen worden. cit. Latif et al. [2012].
Die Art Nuttalliella namaqua kombiniert in ihrer inneren Morphologie bestimmte Eigenschaften der Argasidae und der Ixodidae; sie besitzt aber auch Merkmale, die nur den Nuttalliellidae in einzigartiger Weise zugeordnet sind. Vergleicht man die innere Morphologie einer weiblichen Zecke dieser Art mit der der Familien Argasidae und Ixodidae, so kann man das Folgende feststellen: Die Anzahl der Magenlappen und deren Anordnung sind in allen drei Familien gleich. Allerdings fehlen den Nuttallielliden die umfangreichen Caecalunterteilungen der Argasiden und die lange Caecalschlinge der Ixodiden. Der rektale Sack ist wie bei den Ixodiden nicht gelappt. Der Verlauf der malpighischen Gefäße zwischen den Organen des Körpers gleicht dem der Argasiden. Die transversale Lage des Eierstocks, der zwei-lappige Uterus und die Teilung der Scheide in einen zervikalen und einen vestibularen Teil sind Merkmale der Argasiden. Jedoch finden sich auch Merkmale der Ixodiden, wie zB der den Uterus und die zervikale Vagina verbindende Schlauch und das Verschluss zwischen den Teilen der Vagina. Die sieben Drüsentaschen und deren Anordnung im Gené'sche Organ sind hingegen ausschließliche Merkmale der Nuttalliellidae. Ein dorsaler und vier ventrale Tracheenkanäle, die aus dem Atrium des Spirakulums heraus führen und sich zu etlichen Tracheen verzweigen, sind ⇒ morphologische Merkmale der Argasiden. cit. El Shoura et al. [1984]. Das Hyposthom der Nuttallielliden ist reduziert, apikal abgerundet mit 10-15 Zähnchen in zwei schlecht erkennbaren Reihen.
Die Nuttalliellidae werden wegen dieser Kombination an Merkmalen traditionell als Übergangstaxon, als "missing link", zwischen die beiden großen anderen Familien gestellt. Dabei ist anzumerken, dass die Larve Merkmale besitzt, die weder bei den Schild- noch bei den Lederzecken auftreten: Sie hat Poren auf der ⇒ Dorsalseite der Beine und eine gezähnte Analplatte. Charakteristisch für die Männchen sind das mehr oder minder rechteckige Pseudoscutum, das sich über den größten Teil des Rückens erstreckt, ein Auswuchs auf den ⇒ Chelizeren, der eine einzigartige, stabartige Struktur ähnlich einem ⇒ Spermatodactyl bei anderen Milben bildet, und eine ⇒ mediane Verlängerung des zweiten Pedipalpensegm- eine große, ventrale Mulde zur Beherbergung des vierten Segments bildet. Die Larven und die Nymphen sehen den Weibchen ähnlich. cit. Latif et al. [2012].
Man kann diese Tiere durchaus auch als lebende Fossilien (living fossil), als Überbleibsel aus einer Zeit vor der Trennung von Schild- und Lederzecken betrachten. In diesem Falle besitzt allerdings die Theorie, dass sich die Zecken im Karoo-Basin in Südafrika vor etwa 320 Mill Jahren entwickelt haben, eine hohe Wahrscheinlich keit. cit. Mans et al. [2012]. Als Schädling oder Lästling des Menschen spielen diese Zecken keine Rolle. cit. Keirans et al. [1976].
Nuttalliellidae (1 Gattung, 1 Art)
Die Familie Deinocrotonidae Peñalver, Arillo, Anderson et Pérez-de la Fuente, 2017

Abb. VI.8: Vermuteter Habitus von Deinocroton draculi.
© verändert: Peñalver et al. [2017].
Die Familie Deinocrotonidae ist monotypisch und nur fossil überliefert: Die einzige bisher bekannt gewordene Art Deinocroton draculi Peñalver, Arillo, Anderson et Pérez-de la Fuente, 2017 wurde aus 99 Millionen Jahre altem Burmesischen Bernstein beschrieben. Derzeit wird diese Art als nidikoler Generalist angesehen, der als Adulttier bevorzugt an gefiederten und Nester-bewohnenden Dinosaurieren parasitierte. Die Deinocrotonidae weisen eine Reihe außergewöhnlicher, vermutlich abgeleiteter Merkmale auf: Die Integumentstruktur, die Palpenmorphologie und die Form der Präanalfurche. Die diskontinuierliche Genitalfurche ist ein einzigartiges Merkmal innerhalb der Zecken. Das subterminal gelegene Hypostom und das Vorhandensein eines Pseudoscutums deuten auf eine enge Verwandtschaft zwischen den Deinocrotonidae und den Nuttalliellidae hin, es handelt sich um Schwesterfamilien.
Abgesehen von den oben genannten, die Familie charakterisierenden Merkmalen unterscheidet sich die neue Familie von den Nuttallielliden durch folgende Merkmale:
- Die Oberfläche des Pseudoscutums war wahrscheinlich punktiert, bei den Nuttallielliden ist sie mit einer netzartigen Struktur versehen,
- das Pseudoscutum erreicht bei den Männchen den vorderen Rand des Dorsums,
- Zervikalfurchen sind vorhanden,
- das Capitulum liegt nicht seitlich der Coxae I,
- die Basis capituli ist einfach gestaltet und mit einer glatten Oberfläche versehen,
- Cornua sind nicht vorhanden,
- der Genitalbereich ist glatt, bei den bei den Nuttallielliden ist er unregelmäßig gestreiften,
- anteroventral gibt es einen eingesenkten Bereich in einer postgenitalen Lage, bei den Nuttallielliden in prägenitaler Lage,
- alle Coxen tragen Dornen in Reihen,
- die Beingelenke sind nicht von einem Kugel- und Hülse-Typ, zumindest nicht wie die der Nuttallielliden,
- die proximale Grube des Haller'schen Organs ist vollständig offen,
- die Spirakeln unterscheiden sich in ihrer Morphologie und Größe,
- die Präanal-Furche unterscheidet sich in mikroskopischen Details: Sie hat glatte Ränder, während ihr hinterer und vorderer Rand bei den Nuttallielliden mit integumentalen Zähnchen versehen ist.
cit. Peñalver et al. [2017].
Deinocrotonidae (1 Gattung, 1 Art; nur fossil)
VI.5 Die Unterschiede zwischen den Schild- und den Lederzecken
Die wesentlichen Unterschiede zwischen den beiden rezent existierenden und biologisch bedeutenden Zeckengattungen werden in der folgenden Tabelle zusammengefasst:
| Schildzecken - Ixodidae | Lederzecken - Argasidae |
|---|---|
| Die Cuticula (Körperhülle) ist relativ hart. | Die Cuticula wirkt ledrig, an kleinen warzenförmigen Strukturen setzen im Körperinneren die Muskeln an. |
| Ein Scutum (Schild) aus besonders starrer Cuticula bedeckt den gesamten Rücken der Männchen und einen Teil des Rückens der Weibchen, der Nymphen und der Larven. Das Scutum wird beim Saugakt nicht gedehnt. | Ein Schild ist nicht vorhanden. Männchen und Weibchen unterscheiden sich in dieser Hinsicht äußerlich kaum. |
| Die Mundwerkzeuge ragen über den vorderen Rand der Zecke hinaus, sind also von oben sichtbar. | Die Mundwerkzeuge sind nur bei den Larven von oben sichtbar, bei den anderen Stadien liegen sie auf der Bauchseite des Tieres. |
| Die Tracheenöffnungen (Stigmata) liegen hinter den Coxen des vierten, des letzten Beinpaares. | Die Tracheenöffnungen liegen ventro-lateral neben den Coxen des dritten Beinpaares. |
| Alle Entwicklungsstadien saugen nur einmal, aber meist mehrere Tage lang Blut. | Die Nymphen und Adulten saugen mehrmals, einige Minuten bis zu wenigen Stunden lang Blut. |
| Die Tiere durchlaufen meist nur ein Nymphenstadium. | Die Tiere durchlaufen meist zwei, manchmal bis zu acht Nymphenstadien. |
| Die Männchen sterben nach der Begattung, die Weibchen nach der Eiablage. | Jedes Adulttier kann sich mehrmals paaren. Begattete Weibchen legen nach jeder Blutmahlzeit Eier. |
| Die meisten Schildzecken Österreichs leben im Freien. Jedes Zeckenindividuum befällt in arttypischer Weise in seinem Leben zwei oder, in den meisten Fällen, drei Wirtsorganismen. | In Österreich leben Lederzecken ⇒ synanthrop, meist in Ställen und Dachböden. Jedes Zeckenindividuum befällt mehrere bis zahlreiche Wirtsindividuen. |
| Rücklauf des Plasmas aus der Blutmahlzeit via Speicheldrüse. | Rücklauf des Plasmas aus der Blutmahlzeit via Coxaldrüse. |
| Pulvillus an den Klauen vorhanden. | Pulvillus an den Klauen der Nymphen und Adulten rudimentär oder fehlend. |
Die Familie Argasidae C.L. Koch, 1844, dt: Lederzecken
Die Familie Argasidae besteht gegenwärtig aus 5 Gattungen und 209 Arten. Die Lederzecken kommen in den gemäßigten und kalten Teilen der Welt nur mit einer relativ kleinen Artenzahl vor. Sie bevorzugen heiße und trockene Gebiete. Auch in ⇒ humiden Biotopen bevorzugen sie meist die eher trockenen Nischen. Sie leben meist in Nestern, Ritzen und Spalten in der Nähe von Ruheplätzen von Wirten. Die meisten Argasiden parasitieren auf Vögeln, einige aber auch auf einigen Säugetier- und Reptiliengattungen, darunter auch dem Menschen. Der Saugakt bei den Lederzecken dauert zwei Minuten bis zwei Stunden. Lederzecken können aber auch sehr lange ohne Nahrung auskommen, bis zu zehn Jahren. Die Larven mancher Arten, zB die von Argas reflexus, saugen auch mehrere Tage. Als Krankheitsüberträger werden die Argasiden als weniger wichtig angesehen als die Schildzecken, aber auch sie übertragen einige Infektionserreger. Einige Arten sind von erheblicher Bedeutung in der Veterinärmedizin.
Eine Liste der validen Artnamen aller Lederzecken
Hier eine im Juli 2010 veröffentlichte und damals vollständige, bei Gelegenheit von mir aktualisierte Artenliste de- en der Welt. cit. Camicas et al. [1998], Guglielmone et al. [2009] und Guglielmone et al. [2010].
Argasidae (5 Gattungen, 209 Arten)
Die Familie Ixodidae C.L. Koch, 1844; dt: Schildzecken
Die Familie Ixodidae ist die größte der drei Familien. Sie wird in 14 Gattungen mit zurzeit 743 Arten aufgeteilt. Die Ixodidae bevorzugen im Gegensatz zu den Argasidae ⇒ humide Habitate. Es sind ⇒ obligatorische, permanente ⇒ Ektoparasiten, viele Arten jedoch mehr oder minder ⇒ temporäre Blutsauger. In jedem ihrer Entwicklungsstadien mit Ausnahme des Eies saugen sie nur einmal Blut. Viele Arten sind ausgesprochen ⇒ polyphag, ihre Wirte sind Säugetiere, Vögel oder Reptilien. Dermacentor-Arten saugen sogar an Walrossen und an der Bärenrobbe. Es gibt Schildzecken mit einem drei-wirtigen Entwicklungszyklus, sie wechseln nach jedem Entwicklungsstadium den Wirt. Seltener sind solche mit einem zwei-wirtigen Zyklus, bei denen Larve und Nymphe an demselben Wirtsindividuum saugen, und ein-wirtige. Im Falle von ein-wirtigen Zecken saugen alle Stadien auf demselben Wirtsindividuum ohne es zur Häutung zu verlassen. Der Saugakt dauert lange, meist viele Tage, manchmal verbleibt die Zecke sogar mehrere Monate am Wirt. Bei manchen Schildzecken-Arten saugen die Männchen gar nicht mehr. Während des Saugaktes saugen Schildzecken so viel Blut, dass ihr Körpervolumen sich vervielfacht. Diese Beobachtung war Anlass für Gaius Plinius Secundus Maior (23-79) in seiner Naturalis Historia zu berichten, dass die Zecke von Blut lebe, festsitze und anschwelle, bis sie bei Übersättigung platze.
Eine Liste der validen Artnamen aller Schildzecken
Der Kern der folgenden Liste ist eine im Juli 2010 veröffentlichte und damals vollständige Artenliste der Ixodiden der Welt. Diese wird von mir laufend aktualisiert. Für den gesamten Text ist zu beachten, dass ich die Synonymisierung der Schildzecken weitgehend den Vorschlägen von Guglielmone et Nava [2014] angepasst habe. cit. Camicas et al. [1998], Guglielmone et al. [2009], Guglielmone et al. [2010], Guglielmone et Nava [2014], Nava et al. [2014a], Nava et al. [2014b], Estrada-Peña et al. [2014], Apanaskevich et al. [2013], Apanaskevich et al. [2011] und Apanaskevich et al. [2013].
Ixodidae (14 Gattungen, 743 Arten)
- Margaropus (3 Arten)
- Margaropus reidi Hoogstraal, 1956
- Margaropus wileyi Walker et Laurence, 1973
- Margaropus winthemi Karsch, 1879
- Nosomma (2 Arten)
- Nosomma keralensis Prakasan et Ramani, 2007. Die Originalbeschreibung ist sehr unklar abgefasst.
- Nosomma monstrosum (Nuttall et Warburton, 1908). Dies ist die Typus-Spezies der ehemaligen monotypischen Gattung Nosomma; sie hieß ursprünglich Hyalomma monstrosum.
- Rhipicentor (2 Arten)
- Rhipicentor bicornis Nuttall et Warburton, 1908
- Rhipicentor nuttalli Cooper et Robinson, 1908
- Rhipicephalus (85 Arten - einschließlich von fünf Arten aus dem Genus Boophilus. Diese sind nun in das Genus Rhipicephalus integriert worden)
- Rhipicephalus annulatus (Say, 1821) (Boophilus). Dies ist die Typus-Spezies der Gattung Boophilus. Diese Spezies wurde ursprünglich Ixodes annulatus genannt.
- Rhipicephalus appendiculatus Neumann, 1901
- Rhipicephalus aquatilis Walker, Keirans et Pegram, 1993
- Rhipicephalus armatus Pocock, 1900
- Rhipicephalus arnoldi Theiler et Zumpt, 1949Rhipicephalus aurantiacus Neumann, 1907. Manche Autoren halten diese Art für ein Synonym von R. ziemanni.
- Rhipicephalus australis Fuller, 1899 (Boophilus). Früherer Name: Boophilus microplus (Canestrini, 1888). Einige Autoren geben das Jahr 1887 für die Erstbeschreibung von Rhipicephalus microplus an. Diese Spezies wurde als Haemophysalis micropla [sic] beschrieben. cit. Barker et al. [2014].
- Rhipicephalus bequaerti Zumpt, 1949
- Rhipicephalus bergeoni Morel et Balis, 1976
- Rhipicephalus boueti Morel, 1957
- Rhipicephalus bursa Canestrini et Fanzago, 1878 -> zur Artbeschreibung
- Rhipicephalus camicasi Morel, Mouchet et Rodhain, 1976
- Rhipicephalus capensis Koch, 1844
- Rhipicephalus carnivoralis Walker, 1966
- Rhipicephalus cliffordi Morel, 1965. Diese Art könnte ein Synonym von R. pseudolongus sein.
- Rhipicephalus complanatus Neumann, 1911
- Rhipicephalus compositus Neumann, 1897
- Rhipicephalus congolensis Apanaskevich, Horak et Mulumba-Mfumu, 2013
- Rhipicephalus cuspidatus Neumann, 1906
- Rhipicephalus decoloratus Koch, 1844 (Boophilus)
- Rhipicephalus deltoideus Neumann, 1910
- Rhipicephalus distinctus Bedford, 1932
- Rhipicephalus duttoni Neumann, 1907
- Rhipicephalus dux Dönitz, 1910
- Rhipicephalus evertsi Neumann, 1897
- Rhipicephalus exophthalmos Keirans et Walker, 1993
- Rhipicephalus follis Dönitz, 1910
- Rhipicephalus fulvus Neumann, 1913
- Rhipicephalus geigyi Aeschlimann et Morel, 1965 (Boophilus)
- Rhipicephalus gertrudae Feldman-Muhsam, 1960
- Rhipicephalus glabroscutatum Du Toit, 1941
- Rhipicephalus guilhoni Morel et Vassiliades, 1963
- Rhipicephalus haemaphysaloides Supino, 1897
- Rhipicephalus humeralis Tonelli-Rondelli, 1926
- Rhipicephalus hurti Wilson, 1954
- Rhipicephalus interventus Walker, Pegram et Keirans, 1995
- Rhipicephalus jeanneli Neumann, 1913
- Rhipicephalus kochi Dönitz, 1905
- Rhipicephalus kohlsi (Hoogstraal et Kaiser, 1960) (Boophilus)
- Rhipicephalus leporis Pomerantzev, 1946
- Rhipicephalus longiceps Warburton, 1912
- Rhipicephalus longicoxatus Neumann, 1905
- Rhipicephalus longus Neumann, 1907
- Rhipicephalus lounsburyi Walker, 1990
- Rhipicephalus lunulatus Neumann, 1907
- Rhipicephalus maculatus Neumann, 1901
- Rhipicephalus masseyi Nuttall et Warburton, 1908
- Rhipicephalus microplus (Canestrini, 1888) (Boophilus). Nach der Abtrennung von R. australis ist der Artstatus unklar. Es handelt sich vielleicht um einen Artenkomplex. Einige Autoren geben das Jahr 1887 für die Erstbeschreibung von R. microplus an. Diese Spezies wurde als Haemophysalis micropla [sic] beschrieben.
- Rhipicephalus moucheti Morel, 1965
- Rhipicephalus muehlensi Zumpt, 1943
- Rhipicephalus muhsamae Morel et Vassiliades, 1965
- Rhipicephalus neumanni Walker, 1990
- Rhipicephalus nitens Neumann, 1904
- Rhipicephalus oculatus Neumann, 1901
- Rhipicephalus oreotragi Walker et Horak, 2000
- Rhipicephalus pilans Schulze, 1935
- Rhipicephalus planus Neumann, 1907
- Rhipicephalus praetextatus Gerstäcker, 1873
- Rhipicephalus pravus Dönitz, 1910
- Rhipicephalus pseudolongus Santos Dias, 1953. Manche Autoren halten diese Spezies für ein Synonym von R. longus.
- Rhipicephalus pulchellus (Gerstäcker, 1873)
- Rhipicephalus pumilio Schulze, 1935
- Rhipicephalus punctatus Warburton, 1912
- Rhipicephalus pusillus Gil Collado, 1936
- Rhipicephalus ramachandrai Dhanda, 1966
- Rhipicephalus rossicus Yakimov et Kol-Yakimova, 1911
- Rhipicephalus sanguineus (Latreille, 1806). Das ist die Typus-Spezies des Genus Rhipicephalus; es wurde ursprünglich Ixodes sanguineus genannt -> zur Artbeschreibung
- Rhipicephalus scalpturatus Santos Dias, 1959
- Rhipicephalus schulzei Olenev, 1929
- Rhipicephalus sculptus Warburton, 1912
- Rhipicephalus senegalensis Koch, 1844
- Rhipicephalus serranoi Santos Dias, 1950. Manche Autoren halten diese Spezies für ein Synonym von R. punctatus.
- Rhipicephalus simpsoni Nuttall, 1910
- Rhipicephalus simus Koch, 1844
- Rhipicephalus sulcatus Neumann, 1908
- Rhipicephalus supertritus Neumann, 1907
- Rhipicephalus tetracornus Kitaoka et Suzuki, 1983
- Rhipicephalus theileri Bedford et Hewitt, 1925
- Rhipicephalus tricuspis Dönitz, 1906
- Rhipicephalus turanicus Pomerantzev, 1940. Im Westen wird meist das Jahr 1936 angegeben.
- Rhipicephalus walkerae Horak, Apanaskevich et Kariuki, 2013
- Rhipicephalus warburtoni Walker et Horak, 2000
- Rhipicephalus zambeziensis Walker, Norval et Corwin, 1981
- Rhipicephalus ziemanni Neumann, 1904
- Rhipicephalus zumpti Santos Dias, 1950